远去的犀牛
作者: 吴飞翔

反刍动物是哺乳动物演化史上非常成功的类群——凭借着复杂的消化系统和强大的吸收能力,它们成功逆袭,最终代替了新生代早期风光无限的奇蹄动物。这种此消彼长的背后,是气候环境变化这只看不见的推手,它改变了地表的植被类型,悄然决定了这两类动物的命运。而经历了从热带低地到高寒冰缘的青藏高原,这种演替显得更加彻底。
在青藏高原的现代哺乳动物区系中,偶蹄类种类繁多,包括野猪、林麝、黑麝、高山麝、喜马拉雅麝、毛冠鹿、林麂、赤麂、水鹿、梅花鹿、白唇鹿、马鹿、狍、印度野牛、野牦牛、野水牛、藏原羚、普氏原羚、鹅喉羚、藏羚羊、羚牛、斑羚、赤斑羚、鬣羚、喜马拉雅塔尔羊、岩羊和盘羊,共27种,其中绝大部分是反刍动物,而奇蹄类却仅有藏野驴(马科)1种。但在数千万年前,这里的奇蹄类动物却强势得多。
在5600万~3400万年前的始新世,奇蹄类(现生成员包括马科、貘科和犀科)是地球上主要的有蹄类动物,那时候的奇蹄动物还有已灭绝的雷兽和爪兽。从已有的证据看,除了貘科,马科和犀科都曾在青藏地区生存过。其中犀科动物的种类最多样,存在的时间也最长。从地质时代由新到老,分别有披毛犀、独角犀、大唇犀和近无角犀。
实际上,除了上述几种犀牛,在高原面上我们还在等待另外一类犀牛的出现,那就是地球历史上存在过的最大的陆生哺乳动物——巨犀(Paraceratheriidae)。它们站立时肩高可达5米,头颈伸直可至7米高,体重可达24吨,相当于4头最大的非洲象的体重总和。这种庞然大物在始新世中期(距今约4200万年)起源于蒙古,到中新世早期(距今约2100万年)灭绝。在2000万年的历史里,巨犀纵横欧亚,特别是在渐新世时期,它们穿过尚未整体抬升的青藏地区,往来于亚洲腹地和南亚次大陆之间。从化石记录来看,青藏高原南北两侧的巨犀近亲已经就位,就等高原这最后一块巨犀“拼图”了。
在青藏高原已有的犀类化石记录中,西藏披毛犀是披毛犀家族最早、最原始的成员,它们的后代在冰期来临时走出西藏,成为冰期大型哺乳动物中的代表物种。
大唇犀(Chilotherium)和今天的犀牛不同,雌雄个体都不长角,下颌前端向侧面强烈扩展,这个位置还长有两枚锋利的巨大獠牙,而上颌前部没有门齿,从解剖结构判断可能存在膨大的“上唇”。大唇犀类的祖先可能在中中新世生活于南亚的西瓦立克,随后迁徙到中国、中东和欧洲。晚中新世早期,无角犀族的大唇犀开始出现,随后个体数量快速增加,成为当时动物群中的优势类群。在中国不仅广泛分布于西北地区,也曾来到青藏高原。晚中新世晚期是中国地质历史上犀科动物的全盛时期,大唇犀广布于整个欧亚大陆。随着中新世的结束,中国的犀科动物由盛而衰,高原之上大唇犀最终也被后起之秀披毛犀所取代。
在青藏高原已有的犀类化石记录中,西藏披毛犀是披毛犀家族最早、最原始的成员,它们的后代在冰期来临时走出西藏,成为冰期大型哺乳动物中的代表物种。
大唇犀的化石在西藏两个地点有过发现,一个在藏北那曲地区比如县的布隆盆地,另一个在藏南日喀则的吉隆盆地。最早出现大唇犀化石的比如县布隆现今海拔为4560米,化石时代为晚中新世早期,距今约1000万年。这里的大唇犀为唐古拉大唇犀(Chilotherium tanggulaense),与它伴生的动物有西藏三趾马(Hipparion xizangense)、竹鼠、巨鬣狗、后猫、野猫、萨摩麟(一种原始的长颈鹿) 和羚羊等。这些动物中有不少偏好湿热条件的物种,特别是低冠竹鼠,主要栖息在落叶阔叶林中。除了动物化石,植物的孢粉化石和沉积环境分析结果也显示,当时这里森林密布(孢粉化石说明可能长有棕榈),河湖发育,雨量充沛,土壤处于湿热的氧化环境中。这样的环境与今天这里的高山草甸和高寒干燥的气候环境迥然不同。
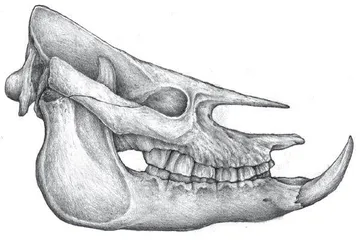
西藏南部吉隆县沃马地点的现代海拔为4384米,一种年代更晚的大唇犀就来自这里,它的时代为晚中新世晚期,年龄经古地磁测定为距今700万年。吉隆的大唇犀名为西藏大唇犀(Chilotherium tibetanensis),它生活在一个多样性也很高的群落里,其他动物有福氏三趾马(Hipparion forstenae)、仓鼠、跳鼠、鼠兔、鬣狗、后麂、古麟(一种原始的长颈鹿)和羚羊等。这些动物的生存环境为森林和草原各占一定比例的疏林地带,古海拔不超过3000米。
另一例犀牛化石来自高原腹地色林错东侧的伦坡拉盆地,那是一块犀牛的肢骨残块。尽管残缺,这块犀牛化石保存的却是一个相当关键的部位,提供了足够的鉴定信息。这是一块前肢的肱骨标本,远端滑车内髁自内向外逐渐收缩,内髁内侧边缘不突起,内上髁发达,向后强烈地伸展,与内髁关节面之间以沟相隔,这些完全符合犀科动物的特点。再与犀科其他属种进行详细对比,根据内髁的形状、内侧副韧带窝和结节的发育程度等特点,可以确定这是近无角犀(Plesiaceratherium)的残骸。
近无角犀是中型到大型的原始无角犀,肢骨细长。在中国,近无角犀此前仅见于两个地点,即山东省临朐县山旺和河北省磁县九龙口。近无角犀的另一个分布地区在欧洲,化石地点有5个。中国产近无角犀的山旺和九龙口动物群的时代是早中新世的山旺期(1800万~1600万年前),而欧洲的化石地点时代也处于早中新世晚期(Orleanian期),与中国化石的时代相当。
当时这件化石对确定伦坡拉盆地这套地层的年代提供了有力的证据。近无角犀化石显示这套地层(丁青组)的上部已经进入中新世。这和化石点附近层段的无脊椎动物化石(介形虫)组合所指示的年代一致。后来,地质学同行在这个剖面上比犀牛化石更低(老)的层位采集了火山灰,测得绝对地质年龄为距今2400万~2300万年,与通过犀牛化石推测的年龄一致(犀牛化石的层位更高,指示的年代更年轻)。

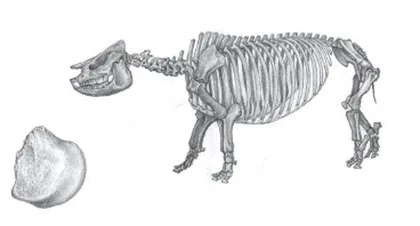
近无角犀的存在对复原当时伦坡拉盆地的古环境也很有意义。近无角犀化石大量出现在山东临朐的山旺动物群中,与三角原古鹿(Palaeomeryx tricornis)和柯氏柄杯鹿(Lagomeryx colberti)都是这个动物群中的优势物种。除了丰富的哺乳动物化石,山旺化石群还含有鱼类、两栖类、爬行类、鸟类、昆虫、大植物、孢粉和藻类等化石,因此可以比较准确地恢复近无角犀的生活环境。
山旺的哺乳动物化石主要为栖息于森林边缘和沼泽地区的类型,尤其是原古鹿、柄杯鹿和各种松鼠等,而在草原生活的类型十分稀少,这说明当时的生态环境是亚热带或暖温带森林。从山旺的植物群来看,其中不少是亚热带常绿或落叶阔叶植物,也显示温暖而湿润的气候。临朐山旺所在的山东半岛中部海拔不超过 1000米,山旺动物群的生态环境表明早中新世时期的海拔应与现代相差不大。近无角犀在欧洲也分布在低海拔地区(海拔低于1000米),享受着早中新世时期(距今1800万~1650万年前)温暖湿润的气候。由此可以看出,近无角犀是一种生活于亚热带或暖温带森林里的动物,偏爱温暖湿润的环境。
那在近无角犀的时代,伦坡拉盆地的海拔有多高呢? 这是一个仍在争论的问题,根据不同证据推测的古高度从1000米左右到4500米不等。如果直接对比,近无角犀在山东和西欧分布区的海拔支持1000米左右的推测值。然而,更合理的推断还要考虑早中新世全球气候背景下动植物分布的上限,因为当时的气候比今天要温暖。
青藏高原南缘喜马拉雅山南坡动植物的分布随着海拔梯度而变化,呈现明显的垂直分带,常绿阔叶林带的分布上限为海拔2500米,气候温暖湿润,年降水量可达2000毫米左右,栖居在此的动物不但种类繁多,而且数量丰富。从动植物组合的特点来看,含近无角犀的山旺动物群以及伦坡拉近无角犀的生存环境都与之相似。
在全球气候背景下,近无角犀生活在早中新世两个变冷事件之间,但温度仍然高于现代,根据氧同位素计算的温度比现代约高4℃。植物垂直带谱的分布与气温直接相关,由于气温直减率为0.6℃/100米,因此,4℃的温度升高可使带谱界线上升约670米,即早中新世时适合近无角犀生活的常绿阔叶林带最高可分布于3170米的海拔处。
从犀科同类的情况来看,现代青藏高原南侧包括尼泊尔在内的南亚地区今天仍有犀科动物分布,印度犀(Rhinoceros unicornis)就生活于喜马拉雅山脚的森林和高草地带。在现生犀牛中,苏门答腊犀(Dicerorhinus sumatrensis)由于身体被毛,因此可以生活在海拔较高的热带森林环境中,栖息地海拔可达1000~1500米。爪哇犀(Rhinoceros sondaicus)最高的分布记录是海拔2000米。这些现代犀牛的生态适宜范围可作为推测近无角犀分布空间的参考,再综合植被分布上限的可能值,考虑早中新世较温暖条件下产生的670米的高差校正,可以推测近无角犀在伦坡拉盆地的栖息地古海拔上限接近3000米。
这样的林地自然不会被犀牛所独享。在找到犀牛化石的那一次考察中,我们还采集到了鸵鸟蛋皮的化石(Struthio sp.),可见鸵鸟也曾在这里繁衍生息。2014年,我们回到这个化石点,在采集的近100袋砂样中,古脊椎所李强研究员筛出了一些小型啮齿动物和爬行动物的牙齿化石。他是研究啮齿动物演化的专家,在阿里札达、昆仑山口、柴达木盆地做了很多啮齿动物化石的研究。除了这些小动物的牙齿以及已经识别出的孢粉化石,我们还采到了双翅目昆虫和挺水型蕨类植物叶片的化石。尽管这些只是当时世界的零星残片,但也够让我们畅想一番 2000万年前藏北腹地的样子。